
Drosera scorpioides, a self-incompatible pygmy Drosera. You need to cross two different clones to get seeds from the flowers.
Self-incompatibility (SI) of flowers is a common theme among plants with about 50% of plant species being afflicted. Self-incompatible plants are not able to produce seeds when its flowers are pollinated from its own flowers or flowers from plants that are genetically the same. Many plants have adaptations to keep individual flowers from physically pollinating themselves but this is something completely different. SI is an active process where "self" pollen tubes are killed in the female flower parts to prevent fertilization. For more information on the mechanisms of self-incompatibility please see the excellent Wikipedia article. Note that only gametophytic self-incompatibility (GSI) and sporophytic self-incompatibility (SSI) are true SI. Heteromorphic "self-incompatibility" is a misnomer. Heteromorphic flowers can be selfed. It has not been determined whether Drosera are GSI or SSI.
Self-incompatibility is found in probably half of the Drosera species. Chen et al. (1997) did a detailed study of self-incompatibility in a selection of Drosera species. The two pygmy SI species they studied, Drosera enebba and Drosera mannii, showed the site of incompatibility to be in the stigma. That is self pollen placed on the stigma started to grow but was killed before it got very far. However the site of incompatibility for the 14 tuberous SI species/subspecies were all over the place: 4 on stigma, 5 in stigma, 1 upper mid-style, 3 mid-style, 1 lower mid-style, 1 placenta, and 1 near ovule. I was astonished to see Drosera auriculata listed as SI in their study. Since they got no seeds one way or another from their source material we cannot be sure it was fertile.
For carnivorous plant enthusiasts, self-incompatibility is a serious problem for the production of seeds if for instance all the plants you have are clones of the same plant. This also presents a serious problem to plant species in nature since SI limits dispersal of a species over long distances. If one seed of an SI species makes it to a new locality, it is essentially doomed not to reproduce while all it takes is one seed for a self-compatible (SC) species to colonize a new locality. This restriction on the ability SI species to establish colonies can result in extinction as continents move, climates change, and local disasters occur.
The decreased ability of SI species to form distant colonies results in most if not all SI Drosera being locally endemic. That is they are found only in small geographic areas. All it takes is a corporate wheat farm to come into production to wipe out a whole species. Whereas self-compatible species tend to be found across larger geographic areas and many are found on different continents and/or islands. An SC species may lose local populations but the species will continue.
So why is self-incompatibility so common among flowering plants? One reason is SI species appear to have a higher rate of speciation. Small SI populations can have as much genetic variation as a large, dispersed SC population. The SI species with small populations in a limited ecological range can quickly develop localized characters taxonomists use to distinguish species. So a human studying a group of species may distinguish a dozen different types and name them as species. But they all may be able to interbreed when grown together and be fertile and self-incompatible like their parents. This is no different from a single large widespread SC species with multiple ecotypes but no obvious taxonomically distinguishing characters. In other words the count of SI vs. SC species may be biased depending on the definition of "species".
Beyond a count of SI and SC species, there is no doubt that self-incompatibility has been a key factor in the diversification of flowering plants. Presumably because of the high rate of speciation and diversification in self-incompatible species, SI species in the past were more likely to found new orders, families, and genera than self-compatible species.
In plant families where self-incompatibility is found today, it is generally considered to be ancestral to the family. This is because we have no examples of self-incompatibility evolving within families or at any level where we can study that process. We do not know how the two SI systems (SSI and GSI) evolved or how many times each evolved independently. The systems work in very complicated ways and it is hard to see how to get there from anywhere else. In phylogenies where the systems have been studied in sufficient detail they find related genera (e.g. petunias and tomatoes) with the same exact genes involved in SI but once that SI system is lost in a clade, it is not regained. This is also what we see in the Caryophyllales carnivores.
Within the Caryophyllales carnivores we have two examples of SI and thus presume the progenitor of this group was SI. The most obvious example is the numerous Australian Drosera exhibiting SI. The other example is Nepenthes; not Nepenthes itself but the likely mechanism for production of dioecy in polyploids is to start with SI parents. See the Nepenthes phylogeny for more info. For these reasons I traced the red SI line in the figure below from the outgroup to where each clade converted to a non-SI mating system. In most of the cases in Drosera, the resulting self-compatible species also self-pollinate. That is the case with most of the species linked with the green lines. Some species are capable of producing viable seed from self pollen but have evolved mechanical ways to limit self-pollination. Those species are shown linked with blue lines.
Three species are shown with dotted lines indicating some populations of those species are SI while others are SC. Drosera indica, Drosera binata and Drosera peltata are actually multimorphic species or species groups depending on your point of view. The founder of the species was probably SI but individual populations have become SC following SI breakdown. At this point it is not known if SI/SC tracks certain forms of each species or is independent of taxonomic status. Each of these species is widespread and SI is generally limited to populations in Australia and Tasmania.
-- John Brittnacher
January 2010

Drosera falconeri hybrid. D. falconeri is self-incompatible but readily makes hybrids with other woolly sundews. The hybrids are also self-incompatible.

Hybrids of another self-incompatible woolly sundew, Drosera derbyensis.

Female flowers from a population of Drosera indica near Narrabri, New South Wales, Australia. This population appears to have developed sexual dimorphism with functionally female flowers and hermaphroditic flowers. The hermaphroditic flowers have autonomous self-pollination. This would be an interesting population to study the evolution of sexual dimorphism in plants. Photo by Robert Gibson, Carniv. Pl. Newslett. 31(1):22 ( PDF ) .
The following list of SI and SC species is not official. It is based on my experience, correspondence with other growers, postings on-line, and publications. If you have any corrections, please contact John Brittnacher,
.
For some species listed there is confusion about whether the species is all or partial self-compatible or self-incompatible. Some of that confusion results from shifting taxonomy but also some populations of the species may have lost self-incompatibility following long-distance dispersal. For instance, Drosera binata is known to be self-incompatible in Australia and Tasmania but self-compatible in New Zealand.
Tuberous Drosera
Self-compatible
D. bulbosa (some populations ?)
D. gracilis
D. gunniana
D. hookeri
D. peltata
D. ramellosa
D. yilgarnensis
Self-incompatible
D. bulbosa (most populations)
D. erythrorhiza
D. gigantea
D. macrantha subsp. macrantha
D. menziesii subsp. basifolia
D. menziesii subsp. menziesii
D. orbiculata
D. porrecta
D. rosulata
D. rupicola
D. stolonifera
D. tubaestylis
Pygmy Drosera
Self-compatible
D. leucostigma
D. nitidula
D. occidentalis
D. omissa (D. ericksoniae)
D. patens (D. nitidula subsp. omissa)
D. pulchella
D. pygmaea
Self-incompatible
D. allantostigma
D. barbigera
D. echinoblastus
D. eneabba
D. mannii
D. meristocaulis (South America)
D. oreopodion
D. paleacea subsp. paleacea
D. paleacea subsp. trichocaulis
D. pedicellaris
D. parvula
D. roseana
D. scorpioides
Woolly Drosera
Self-compatible
D. banksii
Self-incompatible
D. brevicornis
D. broomensis
D. caduca
D. darwinensis
D. derbyensis
D. dilatatopetiolaris
D. falconeri
D. fulva
D. kenneallyi
D. lanata
D. ordensis
D. paradoxa
D. petiolaris
Other Australia/New Zealand Drosera
Self-compatible
D. arcturi
D. adelae (dichogamous)
D. binata (New Zealand, some locations in Australia and Tasmania)
D. burmannii
D. glanduligera
D. hartmeyerorum
D. indica-complex (Australia, Asia, Africa)
D. prolifera (dichogamous)
D. schizandra (dichogamous)
D. spatulata
Self-incompatible
D. binata (most locations Australia, Tasmania)
D. hamiltonii
D. finlaysoniana (some locations in Australia?)
Other Drosera
All self-compatible
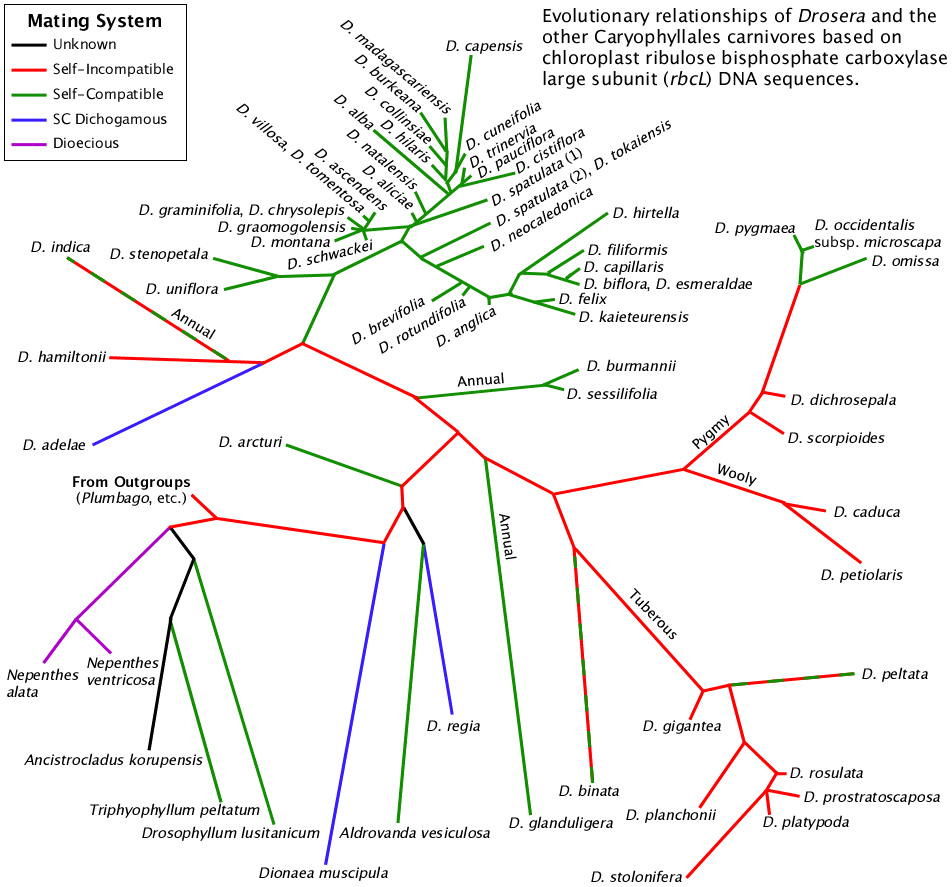
Phylogenetic tree from Drosera evolution page showing the presence of different mating systems in the Caryophyllales carnivores. In self-incompatible plants the female part of the flower kills the pollen with the same genotype as its own to avoid self fertilization. Self-compatible plants are capable of inbreeding. Dichogamous plants have flowers that mature their sexual parts at different times to decrease the probability of inbreeding. Dioecious plants have either male or female flowers and are incapable of inbreeding.
All species are perennial unless noted. The pygmy Drosera perform extensive clonal reproduction via gemmae. Multi-color dotted lines (D. binata and D. peltata) indicate species polymorphic for mating type.
Notes on Drosera self-Incompatibility:
Chen, L., James, S. H. & Stace, H. M. (1997) Self-incompatibility, seed abortion and clonality in the breeding systems of several Western Australian Drosera species (Droseraceae). Austral. J. Bot. 45:191–201. (Detailed study of selected species.)
Lowrie, A. & J. G. Conran. (2007) A revision of the Drosera omissa/D. nitidula complex (Droseraceae) from south-west Western Australia. Taxon 56(2):533–544. (Contains notations as to SI/SC for species complex.)
Gibson, Robert (1999) Drosera arcturi in Tasmania and a comparison with Drosera regia. Carniv. Pl. Newslett. 28(3):76-80 ( PDF ) (Observations on pollination of D. arcturi in nature that are compatible with selfing.)
Gibson, Robert (2002) Drosera indica from Narrabri, New South Wales. Carniv. Pl. Newslett. 31(1):20-24. ( PDF ) (Report on a sexually dimorphic population of what is now named D. finlaysoniana.)
Observations by Greg Bourke and Ivan Snyder indicate D. adelae, D. prolifera and D. schizandra are dichogamous but seed set is poor with selfed plants.
Observations by Andreas Fleischmann indicates D. stenopetala is self-compatible and some D. indica populations are self-incompatible.
Based on numerous observations reported on Internet forums it appears D. hamiltonii is self-incompatible. The plants have large showy flowers typical of SI species and there are no reports of getting seeds from selfed flowers.
Further reading on self-Incompatibility evolution:
Barrett, S. C. H. (2002) Evolution of sex: The evolution of plant sexual diversity. Nature Reviews Genetics 3:274-284.
Igic B., Kohn J. R. (2001) Evolutionary relationships among self-incompatibility RNases. Proceedings of the National Academy of Sciences, USA 98: 13167–13171.
Igic B., L. Bohs, and J. R. Kohn. (2004) Historical inferences from the self-incompatibility locus. New Phytologist 161:97-105.
Igic B., L. Bohs, and J. R. Kohn. (2006) Ancient polymorphism reveals unidirectional breeding system transitions. Proceedings of the National Academy of Sciences 103:1359-1363.
Igic, B., W. A. Smith, K. A. Robertson, B. A. Schaal and J. R. Kohn. (2007) Studies of self-incompatibility in wild tomatoes: I. S-allele diversity in Solanum chilense Dun. (Solanaceae). Heredity 99, 553–561.
Igic, B, R. Lande and J. R. Kohn. (2008) Loss of Self-incompatibility and its Evolutionary Consequences. Int. J. Plant Sci. 169(1):93–104.
Goldberg, E. E., J. R. Kohn, R. Lande, K. A. Robertson, S. A. Smith, B. Igic. (2010) Species Selection Maintains Self-Incompatibility. Science 330:493-495.